Ruffles and Sphincters Control the Spigot of Fresh Blood in the Brain
Quick Links
As the body’s most energy- and oxygen-hungry organ, the brain also happens to be the most dynamic, and it's devilishly complex. How does nutrient-rich blood wend its way through the vast labyrinth of cerebral blood vessels to nourish the neurons that need it most? Two new studies describe elements of neurovascular physiology that make this feat possible. One, published in Nature on February 19 and led by Chenghua Gu at Harvard Medical School, reports that endothelial cells that line arterioles sport myriad inlets, called caveolae, which somehow control the rapid dilation of arterioles in response to neuronal stimulation. The other, published January 20 in Nature Communications and led by Martin Lauritzen of the University of Copenhagen, describes specialized sphincters that control the flow of blood from the brain’s arterioles into its vast capillary beds.
- Indentations called caveolae line the surface of arteriole endothelial cells.
- These caveolae mediate changes in blood flow in response to neuronal activity.
- Specialized sphincters control the flow of blood from arterioles to capillaries in the brain.
"The two papers, by Chow et al and Grubb et. al, significantly advance our understanding of neurovascular physiology and blood flow control," Lei Tong and Jamie Grutzendler of Yale University Medical School, wrote to Alzforum.
Neuronal activity triggers a boost in the regional supply of oxygenated blood within milliseconds. Known as neurovascular coupling, this phenomenon facilitates the coordinated activity of neural networks throughout the brain. It is also the basis of functional brain imaging. While the exact mechanisms involved remain unclear, and controversial, the conventional view is that in response to stimuli, neurons and astrocytes release factors that relax smooth muscle cells surrounding arterioles, thus dilating the vessels and upping the flow of oxygenated blood into the capillary beds connected to them (Jun 2015 news; Iadecola, 2017). However, other reports implicate the endothelial cells lining the arterioles in modulating neurovascular coupling (Chen et al., 2014; Longden et al., 2017).
In their Nature paper, first authors Brian Chow and Vicente Nuñez cast arteriolar endothelial cells as key players, as opposed to mere bystanders, in neurovascular coupling. The researchers observed that, in contrast to the smooth lumen of brain capillaries, the inner surface of arterioles was ruffled, marked by numerous inlets called caveolae.
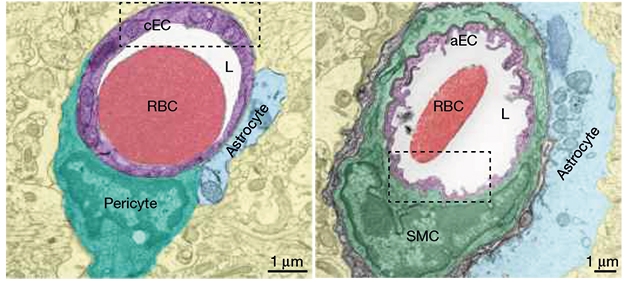
Ruffled Arterioles. Compared with the smooth surface of the capillary lumen (left, purple), the endothelial cells of arterioles (right, purple) were covered with caveolae. [Courtesy of Chow et al., Nature, 2020.]
To investigate the potential role of these caveolae in neurovascular coupling, the researchers used two-photon imaging to peer through cranial windows. They simultaneously measured neuronal activity and dilation of arterioles. Specifically, they brushed a mouse’s whiskers, then assessed the resulting neuronal and vascular responses in the barrel cortex, a region of the rodent somatosensory cortex. As expected, whisker brushing evoked a rapid uptick in neuronal calcium signaling in the region, then local arterioles dilated and red blood cells in downstream capillaries accelerated.
Using conditional knockout mice lacking caveolin-1, a key element of caveolae, the researchers confirmed that caveolae in arteriolar endothelial cells, but not in the smooth muscle cells encircling the vessels, were required for neurovascular coupling. They also found that arteriolar caveolae influenced neurovascular coupling independently of nitric oxide—a critical vasodilatory factor.
Finally, Chow and Nuñez reported that the reason arterioles have caveolae, while capillaries don’t, comes down to expression of MFSD2A. Previously, Gu had reported that capillaries express high levels of this protein, which inhibits caveolae and maintains the integrity of the blood-brain barrier (Ben-Zvi et al., 2014). They found that arterioles express scant MFSD2A. When they overexpressed the protein in arterioles, caveolae largely disappeared, along with neurovascular coupling.
How do caveolae facilitate neurovascular coupling? That’s still unclear, and piecing together the signals among neurons, capillaries, and arterioles is a major focus in her lab, Gu said. She proposed that the caveolae cluster critical ion channels necessary to respond to incoming signals from capillaries, which release factors in response to neuronal stimulation. The arteriolar endothelial cells then signal to smooth muscle cells on the outside of the vessel, triggering dilation.
This proposed chain of events stands in contrast to the conventional “outside-in” model, which posits that vasodilatory factors released from neurons directly relax smooth muscle cells on the outside of the vessel. “In our model, arteriolar endothelial cells play an active role in neurovascular coupling,” Gu said. Neurovascular coupling is disrupted in aging and neurodegeneration, but whether this is a cause or consequence of disease, and how arteriolar caveolae come into play, remain to be deciphered.
In a joint comment to Alzforum, Andrew Yang and Tony Wyss-Coray of Stanford University articulated questions stimulated by these findings. Scientists have reported changes in expression of multiple genes, including caveolin-1, in brain endothelial cells with age and disease, implicating them in the diminution of neurovascular coupling (Nov 2019 news; May 2019 news). “The current study provides new impetus for studying the fascinating complexity of the brain vasculature and will hopefully pave the way toward a better understanding of how this structure degenerates with age and disease,” they wrote.
“[The study] also raises the question whether a similar mechanism operates at the level of brain capillaries, since some recent studies have suggested that capillary dilation precedes arteriolar dilation during neurovascular coupling,” noted Berislav Zlokovic of the University of Southern California in Los Angeles.
In a joint comment to Alzforum, Thomas Pfeiffer, Chanawee Hirunpattarasilp, and David Attwell of University College London made a similar point, noting that caveolin-1 expression on arterioles greatly influenced blood flow in capillaries, more than would be predicted by arteriolar dilation alone. “This raises the question of whether the change of neurovascular coupling that they see on deleting caveolin-1 is not, as one might expect, occurring at the arteriole smooth muscle adjacent to the endothelial cells where the caveolae are being suppressed, but somehow instead at downstream capillary pericytes,” they wrote. Attwell and others have proposed that cerebral blood flow slows in the AD brain because contractile pericytes constrict capillaries, not because smooth muscle cells encircling arterioles squeeze those vessels (Jun 2019 news).
First author Søren Grubb and colleagues addressed connections between arterioles and capillaries in their Nature Communications paper. As their name suggests, penetrating arterioles flow deep into the brain. They supply freshly oxygenated blood to numerous capillaries that branch off along the way. These capillaries, in turn, deliver oxygen and glucose to the neurons and other cells that need it. But how is this process managed in such a way that the capillaries receive an adequate, but not overwhelming, supply of arterial blood? Grubb and colleagues reported that specialized sphincters, positioned on capillaries just as they branch off of arterioles, tightly control the flow of blood into capillary beds.
The researchers used two-photon microscopy to visualize penetrating arterioles in mice that expressed a red fluorescent protein under the control of NG2 promoter, which is expressed in mural cells that surround vessels, including smooth muscle cells and pericytes. They spotted numerous pinched regions of capillaries, each surrounded by a single mural cell, at numerous branch points along penetrating arterioles. Many of these sphincters were followed by a distended region, or bulb. Out of 108 penetrating arterioles with 602 branches that they examined, 433 (72 percent) contained at least one of these sphincters. Most resided on first-order capillaries, at the proximal branch point from arterioles, and were bolstered by a structural skeleton of collagen and elastin that helped maintain the indentation.

Vascular Bottlenecks. Precapillary sphincters (boxed region) appeared at branch points between arterioles and capillaries. The indentations were surrounded by a single mural cell (red). [Courtesy of Grubb et al., Nature Communications, 2020.]
These sphincters dilated rapidly in response to whisker stimulation, then constricted for about 20 seconds before returning to their baseline diameter. During stimulation, the sphincter’s relative diameter increased two to three times more than did the diameter of the arteriole and capillary, and the dilation was controlled by the encircling mural cell.
These sphincters dilated rapidly in response to whisker stimulation, then constricted for about 20 seconds before returning to their baseline diameter. During stimulation, the sphincter’s relative diameter increased two to three times more than did the diameter of the arteriole and capillary, and the dilation was controlled by the encircling mural cell.
Based on the morphology of the mural cells and the markers they expressed, Lauritzen said the sphincter-forming cells are most likely giant, contractile pericytes, as opposed to smooth muscle cells. The sphincters shortened during dilation, and elongated during constriction. Using computational modeling, the researchers concluded that the sphincters serve as vascular bottlenecks, shielding tiny capillaries from relatively high blood pressure in arterioles, while easing in just the right amount of oxygenated blood in response to stimulation. The sphincters also ensured an even distribution of blood along the cerebrovascular tree.
Manning the Flood Gates. A precapillary sphincter widens in response to stimulation of nearby neurons. [Courtesy of Grubb et al., Nature Communications, 2020.]
Lauritzen told Alzforum that the sphincters not only ensure adequate brain perfusion and neurovascular coupling, but also protect the brain from the heart’s pulse. Lauritzen believes that, as blood vessels stiffen with age, they become less adept at attenuating the rush of blood into the brain. To compensate, precapillary sphincters may clinch up, he proposed. This would lead to inadequate perfusion of the brain, and also cause microhemorrhages in backed-up arterioles. Both these problems occur with age and during neurodegeneration, he noted.
How does neuronal stimulation control the sphincters? The signaling pathway remains unclear, but Pfeiffer, Hirunpattarasilp, and Attwell commented that conceivably, pericytes or endothelial cells on capillaries downstream of the sphincter-containing vessel may sense neuronal activity and send a signal back to the sphincter to alter blood flow. “It will be exciting to determine whether, in AD, constriction of the pericytes forming the sphincters has a larger effect on cerebral blood flow than constriction of contractile pericytes that are not forming sphincters, or whether the sphincter-fed vessels are relatively protected from constriction,” they wrote.
“Apart from preventing cerebrovascular complications related to high blood pressure, the sphincters may have a role in smooth vasomotion and the motive force for drainage of interstitial fluid from the brain,” commented Roxana Carare of the University of Southampton, U.K. (Carare et al., 2020). The molecular pathways of neurovascular coupling and dysregulation may contai therapeutic targets for stroke or subarachnoid hemorrhage, Carare noted. —Jessica Shugart.
References
News Citations
- Smooth Muscle Cells, Not Pericytes, Control Brain Blood Flow
- Scientists Discover a Common Distress Signal in the Blood-Brain Barrier
- Paper Alert: VCAM1 Opens the Door to Brain Aging
- Aβ Acts Through Pericytes to Throttle Brain Blood Flow
Paper Citations
- Iadecola C. The Neurovascular Unit Coming of Age: A Journey through Neurovascular Coupling in Health and Disease. Neuron. 2017 Sep 27;96(1):17-42. PubMed.
- Chen BR, Kozberg MG, Bouchard MB, Shaik MA, Hillman EM. A critical role for the vascular endothelium in functional neurovascular coupling in the brain. J Am Heart Assoc. 2014 Jun 12;3(3):e000787. PubMed.
- Longden TA, Dabertrand F, Koide M, Gonzales AL, Tykocki NR, Brayden JE, Hill-Eubanks D, Nelson MT. Capillary K+-sensing initiates retrograde hyperpolarization to increase local cerebral blood flow. Nat Neurosci. 2017 May;20(5):717-726. Epub 2017 Mar 20 PubMed.
- Ben-Zvi A, Lacoste B, Kur E, Andreone BJ, Mayshar Y, Yan H, Gu C. Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature. 2014 May 22;509(7501):507-11. Epub 2014 May 14 PubMed.
Further Reading
Papers
- Vanlandewijck M, He L, Mäe MA, Andrae J, Ando K, Del Gaudio F, Nahar K, Lebouvier T, Laviña B, Gouveia L, Sun Y, Raschperger E, Räsänen M, Zarb Y, Mochizuki N, Keller A, Lendahl U, Betsholtz C. A molecular atlas of cell types and zonation in the brain vasculature. Nature. 2018 Feb 14; PubMed.
Primary Papers
- Chow BW, Nuñez V, Kaplan L, Granger AJ, Bistrong K, Zucker HL, Kumar P, Sabatini BL, Gu C. Caveolae in CNS arterioles mediate neurovascular coupling. Nature. 2020 Mar;579(7797):106-110. Epub 2020 Feb 19 PubMed.
- Grubb S, Cai C, Hald BO, Khennouf L, Murmu RP, Jensen AG, Fordsmann J, Zambach S, Lauritzen M. Precapillary sphincters maintain perfusion in the cerebral cortex. Nat Commun. 2020 Jan 20;11(1):395. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University College London - David Attwell Lab
University College London
Cerebral blood flow (CBF) and the increase in CBF evoked by neuronal activity are reduced early in Alzheimer’s disease (AD) (Iturria-Medina et al., 2016), with the decrease in CBF exceeding 40 percent in affected areas (Asllani et al., 2008). In humans developing AD, and in an AD mouse model, contractile pericytes constrict cortical capillaries sufficiently to generate these CBF decreases (Nortley et al., 2019), and in the AD mouse model there is no change in arteriole or venule diameter implicating pericyte constriction as the main cause of the CBF decrease (Nortley et al., 2019). Two new papers, one (Grubb et al., 2020) from the Lauritzen group in Copenhagen and one (Chow et al., 2020) from the Gu group at Harvard, now report novel mechanisms for the control of CBF, which may have implications for our understanding of the CBF fall that occurs in AD.
CBF can be regulated by vascular smooth muscle cells around arterioles, or by contractile pericytes on the first four branching orders of capillaries coming off an arteriole (Hall et al., 2014). The Lauritzen group now report (Grubb et al., 2020) a specialized structure—termed a sphincter—present on 28 percent of the first-order capillaries branching off cortical penetrating arterioles. The sphincter consists of a narrowed region of the capillary wrapped by an NG2-expressing contractile cell, which conventionally would be called a pericyte (although the paper is careful to avoid getting involved in the debate about the nomenclature of the contractile cells on capillaries) (Attwell et al., 2016).
The authors found that some of these contractile cells were weakly labelled by the Nissl dye NeuroTrace 500/525. This was previously reported to only label non-contractile pericytes (Damisah et al., 2017), but our own unpublished data confirm labelling by NeuroTrace 500/525 of a substantial proportion of α-smooth muscle actin expressing pericytes (which contracted in response to a thromboxane A2 analogue).
The sphincter has a smaller diameter than the adjacent part of the first-order capillary. The authors propose that this allows the sphincter to more significantly reduce the blood pressure being applied to downstream capillaries, and indeed sphincters are found preferentially in the capillaries coming off the penetrating arteriole near the cortical surface where the blood pressure is higher. Conceivably, pericytes or endothelial cells on capillaries downstream of the sphincter-containing vessel may sense neuronal activity and send a signal back to the sphincter to alter blood flow (Longden et al., 2017). It will be exciting to determine whether, in AD, constriction of the pericytes forming the sphincters has a larger effect on CBF than constriction of contractile pericytes that are not forming sphincters, or whether the sphincter-fed vessels are relatively protected from constriction. Detailed computer modelling, based on reconstructions of the vascular topology (Blinder et al., 2013; Kirts et al., 2020) will probably be needed to assess this.
The Gu group paper (Chow et al., 2020) proposes a completely novel mechanism regulating neurovascular coupling, based on the effect of knocking out the protein caveolin-1 from the endothelial cells forming arteries. Caveolin-1 is present in the membranes of caveolae—indentations of the plasma membrane that have been variously proposed to play a role in mediating transcytosis, regulating endothelial cell Ca2+ signalling, sorting membrane proteins, regulating G protein coupled receptors, anchoring GPI-linked proteins, modulating eNOS activity, mechanosignalling, regulating inflammatory pathways, regulating Akt and Erk signalling, and controlling cell proliferation.
Gu’s group had earlier shown that suppression of caveolae-mediated transcytosis in endothelial cells by the lipid transporter Mfsd2a plays a key role in establishing the blood-brain barrier (Andreone, 2018). Surprisingly, they now report that, although this is true for the endothelial cells forming capillaries, which contain few caveolae, there are a large number of caveolae in the endothelial cells forming arterioles. This suggests that the blood-brain barrier may be weaker for arterioles than for downstream capillaries.
When caveolin-1 was knocked out in arterial endothelial cells, caveolae were suppressed in these endothelial cells, and the 15 percent dilation of arterioles normally evoked by neuronal activity was roughly halved (their Fig. 2c), suggesting a major role for caveolin-1 in regulating neurovascular coupling at the arteriole level. A 15 percent increase of arteriole diameter would be expected (by simplistic use of Poiseuille’s law) to evoke a 1.154 = 1.75-fold decrease in arteriole resistance, and hence a 75 percent increase in flow if arterioles constituted the only significant resistance in the cortical vasculature. (In fact, arterioles comprise only around one-seventh of the total vascular resistance from the pial end of a penetrating arteriole to the pial end of a venul (Blinder et al., 2013), so the predicted increase would in fact be much smaller).
Intriguingly, however, the authors in fact documented a fourfold (300 percent) increase in downstream capillary blood flow (their Fig. 2d), indicating that most of the increase in blood flow evoked by neuronal activity is presumably generated by relaxation of contractile pericytes on downstream capillaries. This idea could be tested by measuring capillary diameters near pericytes. This raises the question of whether the change of neurovascular coupling that they see on deleting caveolin-1 is not, as one might expect, occurring at the arteriole smooth muscle adjacent to the endothelial cells where the caveolae are being suppressed, but somehow instead at downstream capillary pericytes.
The paper does not provide mechanistic investigations of how deletion of caveolin-1 alters neurovascular coupling (other than showing it is independent of eNOS activity). Whether the effects observed occur at the arteriole smooth muscle level or the capillary pericyte level, it will be fascinating to determine whether the caveolae-mediated effect is a novel neurovascular coupling mechanism independent of previously known pathways, as has been previously suggested to exist (Hosford and Gourine, 2019) or whether caveolin-1 deletion somehow modulates the properties of an already known pathway.
The role of the caveolin-regulated pathway in disorders such as AD will also be of great interest, since the caveolin-1 KO mouse has already been proposed (Dudau et al., 2016) as a model of dementia, including AD, and there is debate about the effect of changes in the levels of caveolin 1-3 in AD (Gaudreault et al., 2004; van Helmond et al., 2007; Alsaqati et al., 2017).
References:
Iturria-Medina Y, Sotero RC, Toussaint PJ, Mateos-Pérez JM, Evans AC, Alzheimer’s Disease Neuroimaging Initiative. Early role of vascular dysregulation on late-onset Alzheimer's disease based on multifactorial data-driven analysis. Nat Commun. 2016 Jun 21;7:11934. PubMed.
Asllani I, Habeck C, Scarmeas N, Borogovac A, Brown TR, Stern Y. Multivariate and univariate analysis of continuous arterial spin labeling perfusion MRI in Alzheimer's disease. J Cereb Blood Flow Metab. 2008 Apr;28(4):725-36. PubMed.
Nortley R, Korte N, Izquierdo P, Hirunpattarasilp C, Mishra A, Jaunmuktane Z, Kyrargyri V, Pfeiffer T, Khennouf L, Madry C, Gong H, Richard-Loendt A, Huang W, Saito T, Saido TC, Brandner S, Sethi H, Attwell D. Amyloid β oligomers constrict human capillaries in Alzheimer's disease via signaling to pericytes. Science. 2019 Jul 19;365(6450) Epub 2019 Jun 20 PubMed.
Grubb S, Cai C, Hald BO, Khennouf L, Murmu RP, Jensen AG, Fordsmann J, Zambach S, Lauritzen M. Precapillary sphincters maintain perfusion in the cerebral cortex. Nat Commun. 2020 Jan 20;11(1):395. PubMed.
Hall CN, Reynell C, Gesslein B, Hamilton NB, Mishra A, Sutherland BA, O'Farrell FM, Buchan AM, Lauritzen M, Attwell D. Capillary pericytes regulate cerebral blood flow in health and disease. Nature. 2014 Apr 3;508(7494):55-60. Epub 2014 Mar 26 PubMed.
Attwell D, Mishra A, Hall CN, O'Farrell FM, Dalkara T. What is a pericyte?. J Cereb Blood Flow Metab. 2016 Feb;36(2):451-5. Epub 2015 Oct 14 PubMed.
Damisah EC, Hill RA, Tong L, Murray KN, Grutzendler J. A fluoro-Nissl dye identifies pericytes as distinct vascular mural cells during in vivo brain imaging. Nat Neurosci. 2017 Jul;20(7):1023-1032. Epub 2017 May 15 PubMed.
Longden TA, Dabertrand F, Koide M, Gonzales AL, Tykocki NR, Brayden JE, Hill-Eubanks D, Nelson MT. Capillary K+-sensing initiates retrograde hyperpolarization to increase local cerebral blood flow. Nat Neurosci. 2017 May;20(5):717-726. Epub 2017 Mar 20 PubMed.
Blinder P, Tsai PS, Kaufhold JP, Knutsen PM, Suhl H, Kleinfeld D. The cortical angiome: an interconnected vascular network with noncolumnar patterns of blood flow. Nat Neurosci. 2013 Jun 9; PubMed.
Kirst C, Skriabine S, Vieites-Prado A, Topilko T, Bertin P, Gerschenfeld G, Verny F, Topilko P, Michalski N, Tessier-Lavigne M, Renier N. Mapping the Fine-Scale Organization and Plasticity of the Brain Vasculature. Cell. 2020 Feb 8; PubMed.
Andreone BJ, Chow BW, Tata A, Lacoste B, Ben-Zvi A, Bullock K, Deik AA, Ginty DD, Clish CB, Gu C. Blood-Brain Barrier Permeability Is Regulated by Lipid Transport-Dependent Suppression of Caveolae-Mediated Transcytosis. Neuron. 2017 May 3;94(3):581-594.e5. Epub 2017 Apr 13 PubMed.
Hosford PS, Gourine AV. What is the key mediator of the neurovascular coupling response?. Neurosci Biobehav Rev. 2019 Jan;96:174-181. Epub 2018 Nov 24 PubMed.
Dudau M, Frumosu M, Codrici E, Tanase C, Ana-Maria E. Arguments for caveolin-1 knock-out mice as an Alzheimer’s disease model. Austin Alzheimers J Parkinsons Dis. June 2, 2016
Gaudreault SB, Dea D, Poirier J. Increased caveolin-1 expression in Alzheimer's disease brain. Neurobiol Aging. 2004 Jul;25(6):753-9. PubMed.
van Helmond ZK, Miners JS, Bednall E, Chalmers KA, Zhang Y, Wilcock GK, Love S, Kehoe PG. Caveolin-1 and -2 and their relationship to cerebral amyloid angiopathy in Alzheimer's disease. Neuropathol Appl Neurobiol. 2007 Jun;33(3):317-27. PubMed.
Alsaqati M, Thomas RS, Kidd EJ. Proteins involved in endocytosis are upregulated by ageing in the normal human brain: implications for the development of Alzheimer's disease. J Gerontol A Biol Sci Med Sci. 2017 Jun 26; PubMed.
Stanford University Medical School
UCSF
Brian Chow, Vicente Nuñez, and colleagues from Chenghua Gu’s lab at Harvard reveal the molecular mechanisms for a fascinating and fundamental feature of the brain vasculature: How do blood vessels “know” when neurons and other brain cells need more fuel and blood supply—and how exactly is the process regulated? Do all vascular cells participate in this process or only some, and if so, which ones?
Neuronal activity induces a rapid increase in regional blood flow, referred to as neurovascular coupling. The conventional model posits that this is mediated by neuron-derived vasodilatory factors which directly relax arterial smooth-muscle cells (SMCs). Yet from recent work, it seems brain endothelial cells (ECs) can sense neural activity as well. Perhaps then, vasodilatory signals act first on ECs before being relayed to SMCs.
Chow et al. explore this potential EC-mediated neurovascular coupling by adopting a very elegant approach. They focus on the barrel cortex, where whisker stimulation reliably triggers neural activity, vessel dilation, and blood flow. There, they show that classic NO sensing in smooth muscle cells is insufficient for full neurovascular coupling. Instead, the caveolae enriched in arteriolar ECs are also necessary for effective coupling. Using various cell-type-specific and global gene knockout and overexpression mouse models, they confirm that caveolae in ECs—but not in SMCs—are required for neurovascular coupling. Thus, EC caveolae and general eNOS are necessary for a full coupling response.
These discoveries inspire exciting questions relevant to understanding the biology of the brain vasculature in health, aging, and disease. First, what is the mechanism by which caveolae mediate neurovascular coupling? Are they facilitating transport of specific vasodilatory molecules and what are they? In physiological vasodilation, are there cues to preferentially engage EC caveolae versus the NO pathway—and does this change if not in the barrel cortex?
Second, the authors have previously shown caveolae to mediate relatively nonspecific transport. Caveolae likely transport albumin, fibrinogen, and other possibly detrimental factors into the brain, such as in the context of acute brain trauma. Given the abundance of caveolae in arterioles, what special mechanisms prevent this presumable bulk uptake of blood factors?
Third, how do the documented changes in EC gene expression with aging relate to neurovascular coupling? For example, recent papers document an increase in caveolin expression in aged capillaries. By analogy, can age-upregulated capillary caveolae provide vasodilatory signals to contractile pericytes?
Finally, how does this model change with disease? For example, the vascular accumulation of β-amyloid in cerebral amyloid angiopathy has been correlated with a loss of SMCs. In short, the current study provides new impetus for studying the fascinating complexity of the brain vasculature and hopefully will pave the way towards a better understanding of how this structure degenerates with age and disease.
View all comments by Andrew C. YangUniversity of Southern California
This paper by Gu et al. shows that caveolae in the arterial endothelial cells are required for neurovascular coupling at the level of vascular smooth-muscle cells and arterioles. This is an important and novel finding. It provides new mechanistic insight into the mechanisms of neurovascular coupling.
At the same time, it raises the question whether a similar mechanism operates at the level of brain capillaries, since some recent studies have suggested that capillary dilation precedes arteriolar dilation during neurovascular coupling. This would be an important direction for future studies.
View all comments by Berislav ZlokovicUniversity of Southampton School of Medicine
Grubb et al. provide an excellent structural and functional description analysis of the precapillary sphincters. They express alpha smooth-muscle actin and basement membrane proteins, and their main role appears to be in reducing the perfusion pressure in the capillaries. Apart from preventing cerebrovascular complications related to high blood pressure, the sphincters may have a role in smooth vasomotion and the motive force for drainage of interstitial fluid from the brain (Carare et al., 2020).
Chow et al. demonstrate the presence of caveolae on arterial endothelium. They are essential for neurovascular coupling, whereas caveolae on smooth-muscle cells are not necessary for neurovascular coupling. The major facilitator superfamily domain-containing 2a (Mfsd2a) protein is absent from the caveolae of arterial endothelium, while it is associated with the blood-brain barrier and repair after injury such as subarachnoid hemorrhage.
It will be exciting to decipher the full molecular pathways involved in neurovascular coupling and dysregulation, as they could provide valuable therapeutic targets for stroke or subarachnoid hemorrhage.
References:
Carare RO, Aldea R, Bulters D, Alzetani A, Birch AA, Richardson G, Weller RO. Vasomotion Drives Periarterial Drainage of Aβ from the Brain. Neuron. 2020 Feb 5;105(3):400-401. PubMed.
Yale University
Yale University
The two papers by Chow et al. and Grubb et al. significantly advance our understanding of neurovascular physiology and blood flow control.
Through a combination of elegant in vivo two-photon imaging, molecular manipulations, pharmacology, and electron microscopy, Chow et al. report that neurovascular coupling is mediated by arteriolar endothelium to smooth muscle cell (SMC) interactions via an endothelial caveolae-dependent mechanism.
They show that deletion of Caveolin 1 exclusively in endothelium markedly reduces the amplitude of vasodilation following whisker stimulation. Additional deletion of endothelial nitric oxide synthase (eNOS) almost completely abolished neurovascular coupling. Ectopic expression of MFSD2A, a molecule they previously identified as a suppressor of caveolae formation (Ben-Zvi et al., 2014), also attenuates NVC.
There are several interesting conclusions from this study:
1) The arteriolar endothelium appears to be a central hub through which all other signals (possibly neuronal or astrocytic) need to go to achieve vasomotility. In the absence of caveolae or endothelial eNOS, NVC coupling is essentially abolished.
2) Caveolae are markedly enriched in arteriolar endothelium, but not in capillaries. This is consistent with the observation that the overlying smooth muscle cells (SMCs) but not capillary pericytes are the contractile cells that modulate NVC (Hill et al., 2015; Grutzendler and Nedergaard, 2019; Jun 2015 news).
3) Their observations are consistent with previous studies, mainly in the peripheral vasculature, showing that loss of Caveolins induces endothelial dysfunction and reduces myogenic tone, likely through loss of eNOS signaling (Lian et al., 2019).
4) Their results are complementary to recent work from the Nelson lab showing a role of endothelium in potassium sensing through inward-rectifier K+ (KIR2.1) channels to produce a rapidly propagating retrograde hyperpolarization that causes upstream arteriolar dilation (Longden et al., 2017).
5) Their demonstration of structural and functional regional endothelial differences is consistent with the differential transcriptome of arteriolar and capillary endothelial cells as well as the overlying SMCs and pericytes as observed by single-cell RNA seq analysis (Vanlandewijck et al., 2018) and the differential labeling with the uniquely pericyte-specific (completely excluded from SMCs) Neurotrace 500/525 dye which we previously described (Damisah et al., 2017).
In Grubb et al., the authors implement in vivo two-photon imaging in mice that express a fluorescent reporter in all mural cells (NG2-DsRed) while studying aspects of neurovascular coupling. They report the unexpected finding that in addition to the normal-looking, ring-like SMCs around brain arterioles (Hill et al., 2015), there is a small subpopulation of SMCs that have the appearance of a sphincter with greater degree of contractility, and which are located at the transition between the penetrating arteriole and first-order branches.
They provide some evidence, mostly by modeling, that these sphincter-like SMCs generate the largest changes in resistance of all brain vessels and might be important for protecting vasculature and brain tissue from potentially adverse pressure changes.
This study raises several questions:
1) What is unique about these cells from a cellular and molecular point of view compared to the immediately adjacent SMCs?
2) Are these sphincter-like properties stable over time, or are many of these branchpoint SMCs capable of hypercontractility under certain physiological or pathological circumstances?
3) The authors demonstrate the hypercontractile features of these cells during spreading depolarization, potentially a critical finding in the context of ischemia, migraine, and microvascular occlusions.
In summary, this paper adds significantly to our understanding of the structural and functional diversity of mural cells in the brain.
References:
Ben-Zvi A, Lacoste B, Kur E, Andreone BJ, Mayshar Y, Yan H, Gu C. Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature. 2014 May 22;509(7501):507-11. Epub 2014 May 14 PubMed.
Hill RA, Tong L, Yuan P, Murikinati S, Gupta S, Grutzendler J. Regional Blood Flow in the Normal and Ischemic Brain Is Controlled by Arteriolar Smooth Muscle Cell Contractility and Not by Capillary Pericytes. Neuron. 2015 Jun 23; PubMed.
Grutzendler J, Nedergaard M. Cellular Control of Brain Capillary Blood Flow: In Vivo Imaging Veritas. Trends Neurosci. 2019 Aug;42(8):528-536. Epub 2019 Jun 26 PubMed.
Lian X, Matthaeus C, Kaßmann M, Daumke O, Gollasch M. Pathophysiological Role of Caveolae in Hypertension. Front Med (Lausanne). 2019;6:153. Epub 2019 Jul 10 PubMed.
Longden TA, Dabertrand F, Koide M, Gonzales AL, Tykocki NR, Brayden JE, Hill-Eubanks D, Nelson MT. Capillary K+-sensing initiates retrograde hyperpolarization to increase local cerebral blood flow. Nat Neurosci. 2017 May;20(5):717-726. Epub 2017 Mar 20 PubMed.
Damisah EC, Hill RA, Tong L, Murray KN, Grutzendler J. A fluoro-Nissl dye identifies pericytes as distinct vascular mural cells during in vivo brain imaging. Nat Neurosci. 2017 Jul;20(7):1023-1032. Epub 2017 May 15 PubMed.
Vanlandewijck M, He L, Mäe MA, Andrae J, Ando K, Del Gaudio F, Nahar K, Lebouvier T, Laviña B, Gouveia L, Sun Y, Raschperger E, Räsänen M, Zarb Y, Mochizuki N, Keller A, Lendahl U, Betsholtz C. A molecular atlas of cell types and zonation in the brain vasculature. Nature. 2018 Feb 14; PubMed.
Make a Comment
To make a comment you must login or register.