Research Brief: APP Unraveled—Structural Model Extols ε Cleavage
Quick Links
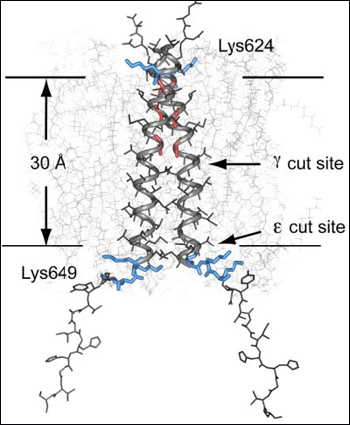
The precise mechanism and significance of APP ε cleavage has been debated since it was discovered some 10 years ago (see ARF related news story). One of the mysteries is that in theory, ε cleavage yields Aβ peptides of 49 amino acids, but in practice, Aβ49 is extremely rare. A paper in this week’s PNAS suggests why this might be. Using solid-state NMR to analyze APP structure, researchers led by Steven Smith at Stony Brook University, New York, confirm that APP exists as a dimer (see Scheuermann et al., 2001) and that it is held together by transmembrane helices. The structure suggests that a transition from helix to random coil right at the edge of the inner cytoplasmic membrane weakens the dimer and allows cleavage by γ-secretase at the ε position, which lies at the polar head of the inner membrane (see figure below).
Structural Data Supports Sequential Cleavage Model of APP Processing
NMR data indicates that APP exists as a dimer held, in part, by interactions between transmembrane helices. Helix-to-random coil transitions at the cytoplasmic side of the domains weakens binding sufficiently to allow ε cleavage by γ-secretase. That cleavage unravels the helical coils further, exposing the APP γ site to proteolysis. Extending the helices by adding three leucines prevents not only ε cleavage, but γ cleavage as well. Image credit: National Academy of Sciences
To test this idea, the researchers extended the helical dimer by adding three leucine residues. This not only eliminated the helix-to-random coil transition near the ε position, but also blocked ε and γ cleavage. The data support the sequential model for γ-secretase action proposed by Yasuo Ihara (see Gu et al., 2001). It posits that APP is first cleaved at the ε position, then at the γ position. According to Smith’s model, that first cleavage would weaken the helices, the protein would become locally unstructured, and the γ-site would become subject to proteolysis. Aβ49 would therefore be lost to Aβ40/42 as the protein is cleaved further.—Tom Fagan
References
News Citations
Paper Citations
- Scheuermann S, Hambsch B, Hesse L, Stumm J, Schmidt C, Beher D, Bayer TA, Beyreuther K, Multhaup G. Homodimerization of amyloid precursor protein and its implication in the amyloidogenic pathway of Alzheimer's disease. J Biol Chem. 2001 Sep 7;276(36):33923-9. PubMed.
- Gu Y, Misonou H, Sato T, Dohmae N, Takio K, Ihara Y. Distinct intramembrane cleavage of the beta-amyloid precursor protein family resembling gamma-secretase-like cleavage of Notch. J Biol Chem. 2001 Sep 21;276(38):35235-8. PubMed.
Further Reading
Papers
- Yu C, Kim SH, Ikeuchi T, Xu H, Gasparini L, Wang R, Sisodia SS. Characterization of a presenilin-mediated amyloid precursor protein carboxyl-terminal fragment gamma. Evidence for distinct mechanisms involved in gamma -secretase processing of the APP and Notch1 transmembrane domains. J Biol Chem. 2001 Nov 23;276(47):43756-60. PubMed.
- Gorman PM, Kim S, Guo M, Melnyk RA, McLaurin J, Fraser PE, Bowie JU, Chakrabartty A. Dimerization of the transmembrane domain of amyloid precursor proteins and familial Alzheimer's disease mutants. BMC Neurosci. 2008;9:17. PubMed.
- Scheuermann S, Hambsch B, Hesse L, Stumm J, Schmidt C, Beher D, Bayer TA, Beyreuther K, Multhaup G. Homodimerization of amyloid precursor protein and its implication in the amyloidogenic pathway of Alzheimer's disease. J Biol Chem. 2001 Sep 7;276(36):33923-9. PubMed.
- Weidemann A, Eggert S, Reinhard FB, Vogel M, Paliga K, Baier G, Masters CL, Beyreuther K, Evin G. A novel epsilon-cleavage within the transmembrane domain of the Alzheimer amyloid precursor protein demonstrates homology with Notch processing. Biochemistry. 2002 Feb 26;41(8):2825-35. PubMed.
Primary Papers
- Sato T, Tang TC, Reubins G, Fei JZ, Fujimoto T, Kienlen-Campard P, Constantinescu SN, Octave JN, Aimoto S, Smith SO. A helix-to-coil transition at the epsilon-cut site in the transmembrane dimer of the amyloid precursor protein is required for proteolysis. Proc Natl Acad Sci U S A. 2009 Feb 3;106(5):1421-6. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Kansas
Sato et al. provide nice evidence that APP residues 618-660 can form a dimer...under conditions used for analysis by NMR. Under these conditions, the APP transmembrane domain appears to assume a helical conformation until the ε cleavage site, at which point there appears to be some disruption of the helix. Whether the APP substrate forms a dimer or not, this disruption of the helical conformation near the ε cleavage site is consistent with a body of data, primarily from Yasuo Ihara's laboratory, that ε cleavage occurs first.
Most of the results in this paper involve spectroscopy, and it is difficult to discern the biological relevance of the findings. The single figure with cellular data (Fig. 5) involves the mutation of three residues near the ε cleavage site, showing that mutation to LLL could prevent proteolysis. One cannot conclude from this experiment, as the authors do, that proteolysis is prevented by extending the transmembrane helix; for example, the LLL insertion extends the hydrophobic transmembrane domain and thus may affect what residues are on the extracellular/lumenal juxtamembrane boundary, which is also known to be important for γ-secretase processing. In addition, the Ihara lab showed essentially the same result by incorporating a stretch of tryptophans in this region (Sato et al., 2005).
References:
Sato T, Tanimura Y, Hirotani N, Saido TC, Morishima-Kawashima M, Ihara Y. Blocking the cleavage at midportion between gamma- and epsilon-sites remarkably suppresses the generation of amyloid beta-protein. FEBS Lett. 2005 May 23;579(13):2907-12. PubMed.
Make a Comment
To make a comment you must login or register.