Newly Identified Microglia Contain Lipid Droplets, Harm Brain
Quick Links
Microglia in the brain assume a dizzying array of states. Now researchers led by Tony Wyss-Coray at Stanford University, Palo Alto, California, describe a new one: lipid droplet-accumulating microglia (LAM). These lipid-stuffed cells resemble the foamy macrophages seen in atherosclerotic lesions. They accumulate in the hippocampus of the aging brain and appear to be bad news, hiking inflammation and reactive oxygen species while having little ability to phagocytose debris. Notably, inflammatory stimuli induce LAM, as do some genetic variants associated with neurodegenerative disease. LAM accumulate in a mouse model of frontotemporal lobar degeneration. The work is described in a preprint posted to bioRxiv (Marschallinger et al., 2019).
- With age, lipid droplet-accumulating microglia (LAM) build up in the hippocampus.
- LAM spew pro-inflammatory cytokines and reactive oxygen species.
- They associate with neuroinflammation and genetic risk factors for neurodegeneration.
Wyss-Coray believes that these microglia promote pathogenesis. “We think they contribute to a damaging pro-inflammatory environment in the brain,” he told Alzforum.
Others agreed that this microglial subtype could shed light on disease. “This work highlights compelling new features of microglia aging, delineating a possible mechanism linking lipid-rich microglia and dementia,” Marco Colonna and Simone Brioschi at Washington University in St. Louis wrote to Alzforum (full comment below). Mathew Blurton-Jones and Christel Claes at the University of California, Irvine, noted that many AD risk genes are involved in lipid processing. “It will be fascinating to determine how the composition of lipids that accumulate in human disease-associated microglia and LAM are influenced by these genes, and to watch how this exciting story continues to evolve,” they wrote (full comment below).
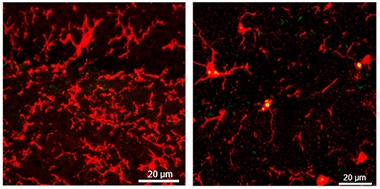
Lipid Droplets Accrue With Age. Lipid droplets (yellow) accumulate in microglia (red) of older mice (right), but not in young mice (left). [Courtesy of Julia Marschallinger and Tony Wyss-Coray.]
Previous studies have identified a smorgasbord of distinct transcriptional profiles delineating subtypes of microglial states (Jul 2018 conference news; Dec 2018 news). A handful of these have been correlated with neurodegenerative disease. These include disease-associated microglia (DAM), which cluster around plaques in mouse models of amyloidosis, and the similar microglial neurodegenerative phenotype (MGnD) found in multiple mouse disease models (Jun 2017 news; Sep 2017 news). A recent study characterized human Alzheimer’s microglia (HAM), which were isolated from AD brain (May 2019 news). It is still unclear how all these types relate to each other and what they do.
Wyss-Coray and colleagues did not set out to do a transcriptional study, rather, they were studying brain aging. While examining hippocampal sections from aged wild-type mice by electron microscopy, first author Julia Marschallinger was struck by the accumulation of lipid droplets inside microglia. These microglia resembled cells first described by Alois Alzheimer, who reported adipose-stuffed glia clustering around amyloid plaques in AD brain more than 100 years ago. Marschallinger and colleagues quantified the phenomenon in mice, finding that more than half the hippocampal microglia in 20-month-old wild-type animals contained from one to three lipid droplets. Droplets were rare in other brain regions, and nearly absent in 3-month-old mice. Lipidomic analysis determined that the droplets consisted of electrically neutral lipids, mainly glycerolipids such as triacylglycerols and diacylglycerols, but almost no cholesterol esters.
To characterize these LAM, the authors isolated microglia from aged mouse hippocampi and sorted out those with high lipid content. Transcriptional profiling revealed 692 genes that were differently expressed between cells with low and high lipid content. In particular, genes involved in the production of reactive oxygen species, lipids, and pro-inflammatory cytokines were up in LAM, while genes responsible for phagocytosis were down. Notably, this transcriptional profile was in many respects the opposite of DAM, which turn up phagocytotic genes.
Functional studies of LAM reinforced these transcriptional findings. When the authors injected myelin debris into aged mouse hippocampus, microglia without lipid droplets engulfed it, but few LAM did. LAM isolated from brain produced more reactive oxygen species (ROS) than did low-lipid microglia, and they secreted higher levels of several pro-inflammatory cytokines such as CCL3, CXCL10, and IL-6.
How do these cells arise? Because many of the LAM genes are regulated by inflammation, the authors speculated they are products of an inflammatory response. To test this, they treated the BV2 mouse microglial line with the inflammatory toxin lipopolysaccharide (LPS). The number of lipid droplets in the BV2 cells increased fivefold. Like LAM in the brain, cells stuffed with lipid droplets poorly phagocytosed a yeast glycan, while producing high levels of ROS. Treating cells with Triacsin C, an inhibitor of glycerolipid synthesis, prevented droplet formation and kept phagocytosis and ROS at normal levels, indicating the lipids were a cause rather than an effect of the microglial transformation. Inflammation also induced LAM in vivo. Intraperitoneally injecting 1 mg/kg LPS into young mice for four days doubled the number of LAM in the hippocampus.
To find genetic factors that could promote LAM, the researchers screened a library of about 2,000 genes involved in cellular stress and protein degradation. They knocked out each gene one by one in BV2 cells, added LPS, then looked for induction of lipid droplets. To quickly find effects, they treated the cells with a lipid marker bound to a phototoxin and blasted them with light, killing off cells with droplets. In this way, they found 112 genes that influenced droplet formation, including several linked to different forms of neurodegenerative disease: SLC33A1, SNX17, VPS35, CLN, NPC2, and GRN. SLC33A1 mutants cause spastic paraplegia, VPS35 variants lead to Parkinson’s, CLN to ceroid lipofuscinosis, and NPC2 mutants to Niemann-Pick disease. GRN produces progranulin, variants of which cause frontotemporal dementia. The authors examined middle-aged GRN knockout mice, which already have inflammation in the brain, and found twice as many LAM in their hippocampi as in age-matched, wild-type mice. GRN knockouts also developed LAM in the thalamus, cortex, and corpus callosum.
“We think this accumulation of lipid droplets is intrinsically linked to neurodegenerative disease,” Wyss-Coray said. In ongoing work, he has found a similar buildup of LAM in mouse models of amyloidosis. He will test whether lowering brain lipids, or eliminating LAM altogether, improves brain health in various mouse models of neurodegeneration.
It is still unknown whether LAM accumulate in Alzheimer’s, but Wyss-Coray and colleagues report finding this cell type in aged human brain. They saw microglia sporting lipid droplets in postmortem hippocampal sections from a cognitively healthy 67-year-old, but few in a 22-year-old. Some previous studies reported finding this microglial phenotype in injured human brain (Lee et al., 1990; Castejon et al., 2005).
Why have more studies not noticed these cells? Wyss-Coray noted that most histological processing of postmortem tissue uses detergents, which removes lipids. In addition, most LAM occur in the hippocampus, so studies of other regions would miss them. The reason for this localization is unclear. Wyss-Coray noted that microglial density is higher in the hippocampus than in other brain regions, as are the functional demands on these cells.
Commenters were also intrigued by this regional specificity. “LAM seem to concentrate in areas subjected to stress,” Lorenza Magno at University College London wrote to Alzforum (full comment below). Blurton-Jones wondered if the presence of LAM in the hippocampus might relate to the turnover of newborn neurons, given the role of microglia in clearing apoptotic cells.
LAM are not the only microglial subtype to accumulate lipids. Julia TCW and Alison Goate at the Icahn School of Medicine at Mount Sinai, New York, recently reported that ApoE4 microglia stockpile lipids; however, these are mostly in the form of free cholesterol, rather than neutral lipids (Aug 2019 news). Wyss-Coray noted that ApoE expression does not change in LAM, reinforcing the idea that ApoE4 microglia represent a different subtype. Meanwhile, researchers in Germany described dysfunctional, lipid-filled microglia in old mice with axon injuries; these cells become choked with myelin debris and crystallized cholesterol (Cantuti-Castelvetri et al., 2018). These microglia are also distinct from LAM, which do not take up myelin, Wyss-Coray said.
Peripheral phagocytes can also accumulate lipids. Ido Amit at the Weizmann Institute of Science, Rehovot, Israel, found that macrophages in mouse and human adipose tissue take up excess cholesterol, helping keep body fat under control. These macrophages express TREM2 and have a similar transcriptional profile to DAM. Amit also dubbed them LAM, but for lipid-associated macrophages. He found that TREM2 knockout mice became fatter on a high-fat diet than did wild-type mice, and developed high cholesterol and insulin resistance. Thus, it seems that these LAM in the periphery help protect against the deleterious consequences of a fatty diet. Amit reported these data at the FENS Brain Conference last May and published in the July 25 Cell (May 2019 conference news).
While these peripheral macrophages and the lipid droplet-accumulating microglia seen in brain are now labeled with the same acronym, Wyss-Coray believes the two cell types have little in common. Not only do the transcriptional profiles differ, but the two types of phagocyte accumulate different lipids as well. Peripheral LAM contain mostly cholesterol and sphingomyelin, unlike the neutral lipids found in brain LAM, Wyss-Coray noted.—Madolyn Bowman Rogers
References
News Citations
- A Delicate Frontier: Human Microglia Focus of Attention at Keystone
- Microglia Reveal Formidable Complexity, Deep Culpability in AD
- Hot DAM: Specific Microglia Engulf Plaques
- ApoE and Trem2 Flip a Microglial Switch in Neurodegenerative Disease
- When It Comes to Alzheimer’s Disease, Do Human Microglia Even Give a DAM?
- ApoE4 Glia Bungle Lipid Processing, Mess with the Matrisome
- Brain Conference Spotlights Transcriptomics, Therapeutic Strategies
Paper Citations
- Marschallinger J, Iram T, Zardeneta M, Lee SE, Lehallier B, Haney MS, Pluvinage JV, Mathur V, Hahn O, Morgens DW, Kim J, Tevini J, Felder TK, Wolinski H, Bertozzi CR, Bassik MC, Aigner L, Wyss-Coray T. Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nat Neurosci. 2020 Feb;23(2):194-208. Epub 2020 Jan 20 PubMed. Correction.
- Lee SC, Moore GR, Golenwsky G, Raine CS. Multiple sclerosis: a role for astroglia in active demyelination suggested by class II MHC expression and ultrastructural study. J Neuropathol Exp Neurol. 1990 Mar;49(2):122-36. PubMed.
- Castejón OJ, Castellano A, Arismendi GJ, Medina Z. The inflammatory reaction in human traumatic oedematous cerebral cortex. J Submicrosc Cytol Pathol. 2005 Apr;37(1):43-52. PubMed.
- Cantuti-Castelvetri L, Fitzner D, Bosch-Queralt M, Weil MT, Su M, Sen P, Ruhwedel T, Mitkovski M, Trendelenburg G, Lütjohann D, Möbius W, Simons M. Defective cholesterol clearance limits remyelination in the aged central nervous system. Science. 2018 Feb 9;359(6376):684-688. Epub 2018 Jan 4 PubMed.
Further Reading
Primary Papers
- Marschallinger J, Iram T, Zardeneta M, Lee SE, Lehallier B, Haney MS, Pluvinage JV, Mathur V, Hahn O, Morgens DW, Kim J, Tevini J, Felder TK, Wolinski H, Bertozzi CR, Bassik MC, Aigner L, Wyss-Coray T. Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nat Neurosci. 2020 Feb;23(2):194-208. Epub 2020 Jan 20 PubMed. Correction.
- Jaitin DA, Adlung L, Thaiss CA, Weiner A, Li B, Descamps H, Lundgren P, Bleriot C, Liu Z, Deczkowska A, Keren-Shaul H, David E, Zmora N, Eldar SM, Lubezky N, Shibolet O, Hill DA, Lazar MA, Colonna M, Ginhoux F, Shapiro H, Elinav E, Amit I. Lipid-Associated Macrophages Control Metabolic Homeostasis in a Trem2-Dependent Manner. Cell. 2019 Jul 25;178(3):686-698.e14. Epub 2019 Jun 27 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Washington University School of Medicine
Washington University in St. Louis
Recent evidence showed that, during aging, microglia accumulate lipids within the phagosomes. Hence, it has been suggested that lipid-rich microglia would less efficiently uptake and degrade cellular debris or protein aggregates, and this may contribute to brain aging and neurodegenerative diseases (Cantuti-Castelvetri et al., 2018; Safaiyan et al., 2016). This recent work by Marschallinger and colleagues has further elucidated this phenomenon, showing that lipid-rich microglia accumulate in the hippocampi of both aging mice and senile humans.
Using RNA sequencing, authors showed that lipid-rich microglia exhibit a different gene expression compared with microglia with low lipid content. Interestingly, most of the upregulated genes are involved in phagosomes maturation and ROS production. The authors named this specific microglial phenotype “Lipid droplet Accumulating Microglia,” or LAM. Using different assays, they demonstrated that LAM exhibit reduced phagocytic capacity and increased production of ROS and pro-inflammatory cytokines. Lastly, the authors showed that brains of progranulin-deficient mice (Gran-/-) exhibit abundant LAMs already during adulthood.
In humans, Grn mutations have been linked to frontotemporal dementia, therefore accumulation of lipid droplets in human microglia may play an important role in the context of neurodegenerative diseases. Overall, this work highlights new compelling features of microglial aging, delineating a possible mechanism linking lipid-rich microglia and dementia. For the future, it will be interesting to determine whether certain Alzheimer’s risk alleles (such as ApoE-ε4 or Trem2 R47H) could exacerbate the LAM formation, thus accelerating brain aging and deposition of Aβ.
References:
Cantuti-Castelvetri L, Fitzner D, Bosch-Queralt M, Weil MT, Su M, Sen P, Ruhwedel T, Mitkovski M, Trendelenburg G, Lütjohann D, Möbius W, Simons M. Defective cholesterol clearance limits remyelination in the aged central nervous system. Science. 2018 Feb 9;359(6376):684-688. Epub 2018 Jan 4 PubMed.
Safaiyan S, Kannaiyan N, Snaidero N, Brioschi S, Biber K, Yona S, Edinger AL, Jung S, Rossner MJ, Simons M. Age-related myelin degradation burdens the clearance function of microglia during aging. Nat Neurosci. 2016 Aug;19(8):995-8. Epub 2016 Jun 13 PubMed.
University of California, Irvine
cclaes@uci.edu
This exciting new study by Marschallinger et al. defines and examines the age-dependent accumulation of lipid droplets within microglia, clearly demonstrating that these lipid-accumulating microglia (LAMs) exhibit a unique transcriptional signature, pro-inflammatory phenotype, and impaired phagocytosis. Interestingly, LAM are far more prevalent in the hippocampus than other brain regions. While the cause of this hippocampal LAM enrichment remains unclear, it is interesting to speculate that this may be associated with the turnover of newborn neurons, given the role of microglia in the clearance of apoptotic cells within neurogenic niches (Fourgeaud et. al., 2016).
Focusing in on the hippocampus, Marschallinger and colleagues use BODIPY, which detects neutral lipids, to quantify and isolate LAM and compare them to microglia with low lipid content. They find that the number of LAM are more than fourfold higher in 20-month-old versus 3-month-old mice, and likewise using an antibody approach provide initial evidence that LAM are also increased in aged human brains. The authors then use their BODIPY FACS approach to examine the transcriptome and functions of LAM, finding significant changes in in phagosome maturation and pro-inflammatory pathways.
While the acronym LAM was also recently introduced by Ido Amit in reference to “lipid-associated macrophages” from white adipose tissue (Jaitin et al., 2019), the LAM described in the current study differ considerably from these adipose LAM. Whereas adipose-LAM exhibit transcriptional profiles that closely mimic signatures of disease-associated microglia (DAM), the age-associated microglial-LAM described in the current study exhibit profiles that are relatively inverse to that of AD-associated DAM and appear to be TREM2-independent. As the presence of amyloid plaques and dystrophic neurites likely alters the lipids that microglia are exposed to, it will be very interesting to learn how the lipid content of DAM differs from that of adipose- and age-associated LAM and whether human DAM and LAM exhibit similar profiles to their murine counterparts.
In this respect, we recently demonstrated considerable differences between human and murine DAM genes in a chimeric AD model, suggesting important species differences exist in these various microglial phenotypes (Hasselman et al., 2019). Interestingly, our ongoing studies have also revealed lipid accumulation and enrichment of many lipid-associated genes in human plaque-associated DAM. Given the growing list of lipid-associated AD risk genes (APOE, TREM2, Clusterin, ABCA7, SORL1, MS4As), it will be fascinating to determine how the composition of lipids that accumulate in human DAMs and LAMs are influenced by these genes and to watch how this exciting story continues to evolve.
References:
Fourgeaud L, Través PG, Tufail Y, Leal-Bailey H, Lew ED, Burrola PG, Callaway P, Zagórska A, Rothlin CV, Nimmerjahn A, Lemke G. TAM receptors regulate multiple features of microglial physiology. Nature. 2016 Apr 14;532(7598):240-244. Epub 2016 Apr 6 PubMed.
Jaitin DA, Adlung L, Thaiss CA, Weiner A, Li B, Descamps H, Lundgren P, Bleriot C, Liu Z, Deczkowska A, Keren-Shaul H, David E, Zmora N, Eldar SM, Lubezky N, Shibolet O, Hill DA, Lazar MA, Colonna M, Ginhoux F, Shapiro H, Elinav E, Amit I. Lipid-Associated Macrophages Control Metabolic Homeostasis in a Trem2-Dependent Manner. Cell. 2019 Jul 25;178(3):686-698.e14. Epub 2019 Jun 27 PubMed.
Hasselmann J, Coburn MA, England W, Figueroa Velez DX, Kiani Shabestari S, Tu CH, McQuade A, Kolahdouzan M, Echeverria K, Claes C, Nakayama T, Azevedo R, Coufal NG, Han CZ, Cummings BJ, Davtyan H, Glass CK, Healy LM, Gandhi SP, Spitale RC, Blurton-Jones M. Development of a Chimeric Model to Study and Manipulate Human Microglia In Vivo. Neuron. 2019 Sep 25;103(6):1016-1033.e10. Epub 2019 Jul 30 PubMed.
ARUK UCL Drug Discovery Institute
Microglia heterogeneity has just started being appreciated. Their diversity likely depends upon intrinsic properties as well as environmental cues within their niche, including specific regional localization. Overlying this diversity, microglia are also dynamic, and may switch between different states in response to an insult. Several phenotypes have been already identified in conditions such as aging or disease. However, the roles of microglia in these different states is far from complete.
Against this backdrop, Marschallinger et al. characterized an additional functional state of microglia that seemingly reflects changes in lipid metabolism and is associated with inflammation, aging, and neurodegeneration. Interestingly, only partial overlap was found with previously described “diseased-microglia” signatures. The so-called “lipid-droplet accumulating microglia” (LAM) constitute a proportion of the microglia population that increases during aging, and seems to be present predominantly in the hippocampus rather than in other brain regions. Is there a regional specificity to the phenotype observed, e.g., does this phenotype reflect regional selectivity to aging? Using a model for frontotemporal dementia (FTD), the authors found additional LAM in the thalamus, a region particularly affected by degeneration in FTD. This suggests that a selective vulnerability might also be linked to the specific disease-context. In any case, LAM seem to concentrate in areas subjected to stress. Whether specific subtypes of microglia within these areas are particularly susceptible to metabolic stress remains to be addressed.
Leukocytes, macrophages, and microglia containing accumulations of lipids upon inflammation have been described before. Lipid droplets have been associated with inflammation and cytokine storage, but also with homeostatic functions, such as storage of fatty acids that could support phagocytosis and proper mitochondria functioning (Nadjar et al., 2018). More broadly, a role for glial cells in maintaining lipid homeostasis has been described, with a recent paper showing that neuronal lipids are transferred to astrocytes, to counteract excitotoxic effects (Ioannou et al., 2019). Although Marschallinger et al. reported that no transcriptional changes have been detected in lipid transporter genes, suggesting that lipid droplet dynamics in microglia might differ compared with astrocytes, some changes could occur at the protein level and on a wider level, as compensatory changes of the entire lipid metabolism. It would be of great interest to assess the crosstalk between different cell types (including neurons, astrocytes, and microglia, but also potentially ependymal cells and endothelial cells lining the blood vessels) that mediate lipid metabolism in the brain.
Through comparison of the transcriptome of LAM with non-lipid-laden microglia, the authors identified a new profile associated with lysosomal, ROS, and NO production genes, with phagocytosis and ROS generation as the highest upregulated pathways. Remarkably, LAM showed severely reduced phagocytosis accompanied by accumulation of lysosomal vesicles. Could droplet numbers increase as a consequence of impaired cell function (e.g., reduced mitochondria functioning and impaired phagocytosis) and reduced lysis? As suggested by the authors, lysosomal defects might underlie the accumulation observed. At the same time, the droplets themselves might be the cause for the altered homeostatic functioning by “sequestering” the lysosomes normally involved in phagocytosis. What is clear is that the phenotype of this cell is detrimental to properly functioning microglia.
One critical question still to be addressed is whether the LAM transcriptional profile can be mapped onto aging and/or diseased microglia in the human brain (such as the recently described HAM signature, Srinivasan et al., 2019). Lipofuscin deposits are often found in human brains, sometimes in association with lipid droplets, however, it is not clear whether they always co-exist and might be the consequence of distinct processes (Shimabukuro et al., 2016). Lipofuscin deposits are a hallmark of a number of degenerative disorders, including age-related macular degeneration (Kauppinen et al, 2017).
Accumulation of lipids is not a unique feature to microglia, however, it seems to be a hallmark of brain aging. GWAS have identified lipid metabolism as an important pathway in AD, and GWAS hits, such as Trem2 and ApoE, are involved in lipid sensing and processing. As mentioned in this paper, Dr. Alzheimer himself described lipid accumulation in the cell cytoplasm of AD brains. This could be an unappreciated feature that characterizes not only AD, but also other neurodegenerative pathologies. Finally, this new field of investigation opens up new questions on the processes occurring during these disorders. For instance, could AD be considered a systemic disease, where a more broadly disrupted metabolism causes locally altered homeostasis, and consequent neurodegeneration due to loss of support from homeostatic functions in the brain?
References:
Nadjar A. Role of metabolic programming in the modulation of microglia phagocytosis by lipids. Prostaglandins Leukot Essent Fatty Acids. 2018 Aug;135:63-73. Epub 2018 Jul 18 PubMed.
Ioannou MS, Jackson J, Sheu SH, Chang CL, Weigel AV, Liu H, Pasolli HA, Xu CS, Pang S, Matthies D, Hess HF, Lippincott-Schwartz J, Liu Z. Neuron-Astrocyte Metabolic Coupling Protects against Activity-Induced Fatty Acid Toxicity. Cell. 2019 May 30;177(6):1522-1535.e14. Epub 2019 May 23 PubMed.
Srinivasan K, Friedman BA, Etxeberria A, Huntley MA, van der Brug MP, Foreman O, Paw JS, Modrusan Z, Beach TG, Serrano GE, Hansen DV. Alzheimer's Patient Microglia Exhibit Enhanced Aging and Unique Transcriptional Activation. Cell Rep. 2020 Jun 30;31(13):107843. PubMed.
Shimabukuro MK, Langhi LG, Cordeiro I, Brito JM, Batista CM, Mattson MP, Mello Coelho Vd. Lipid-laden cells differentially distributed in the aging brain are functionally active and correspond to distinct phenotypes. Sci Rep. 2016 Mar 31;6:23795. PubMed.
Boston University School of Medicine
This is an interesting study that reminds me of Alois Alzheimer’s drawing of Nile red staining, which reveals intracellular lipid droplets (LD) in cells (potentially microglia) around plaques in AD patient brain. Here, Marschallinger and colleagues identified LD-accumulating microglia in aged brain, which led to a defect in phagocytosis, high levels of reactive oxygen species (ROS), and pro-inflammatory phenotypes.
DAM, HAM, and LAM: stage-specific microglia captured from young to old brains?
The authors separated cells containing high vs. low levels of LDs. High-LD microglia (50 percent of those in the aged brain), called LAM, showed unchanged expression of APOE, which is one of the key markers for disease-associated microglia (DAM) polarization, but expression changes in many other genes that were the opposite of that seem in DAM. However, the other half of the aged brain microglia, which are low-LD, may still have a DAM-like signature. The authors’ findings suggest there is an LD-dependent divergence of beneficial and detrimental microglia, which co-exist in aged brain. The relative proportions of these cells probably changes during the time course of aging and are affected by genetic mutations and disease risk factors. Further comparison of the overlap between LAM and human Alzheimer’s microglia (HAM), which represents an enhanced aging phenotype (Galatro et al., 2017; Srinivasan et al., 2019), may reveal age-associated gene-expression changes shared in both mouse and human microglia. Surprisingly, Triacsin C easily reverts the LAM phenotype to normal, but we may need to investigate further how blocking de novo synthesis of lipids affects cellular lipid homeostasis and if it would have potential adverse effects if used as a therapeutic intervention.
Differences between lipid-accumulating microglia and LAM (lipid droplet accumulating microglia)
We need to use the acronym LAM carefully. This study is focused on a cellular organelle, the lipid droplet, but not the entire complement of cellular lipids. There are studies that show microglia accumulate cholesterol after excessive uptake of extracellular lipids or if they express the AD genetic risk factor APOE 4 (Cantuti-Castelvetri et al., 2018; TCW et al., 2019). These lipid-accumulating microglia—i.e., not LAM—possess mainly free (unesterified) cholesterol, which is converted from extracellular lipids or generated through biosynthesis. In LAM, Marschallinger and colleagues found almost no cholesteryl esters in LD of aged microglia and aged hippocampal mouse brain, and their data also suggested that demyelination does not contribute to LD in LAM. Consistent with unaffected levels of cholesteryl ester by APOE4, as measured by gas chromatography-mass spectrometry (TCW et al., 2019), the authors also observed that APOE and other lipid transporters were not significantly regulated in LAM. However, to address cholesterol accumulation in aged microglia, total cholesterol as well as intracellular free cholesterol should be considered.
Regional/cell-type-specific differences in lipid composition of LD
Lipidomic analysis reported in this preprint showed glycerolipids and phospholipids are the major content of LAM. It is also interesting to see the different composition of brain vs. peripheral cells (liver), for example sphingomyelin, which forms a partner with cholesterol in plasma membranes that control cholesterol synthesis, is high in liver cells but not in brain microglia. We should further explore microglia specific lipid regulation.
What’s first, inflammation or lipid accumulation? Whichever comes first, LAM do not phagocytose!
In transcriptome, authors found that lipopolysaccharide (LPS) is the most significant upstream regulator in high LD microglia. Further in vitro validation showed that LPS treatment of BV2 cells and hippocampal microglia in young mice increases lipid droplets, indicating inflammation-induced LD formation. Many other factors can induce LD formation, however, the important finding is that LAM in aged brain do not phagocytose, displaying a unique transcriptomic signature as well as a detrimental phenotype.
References:
Galatro TF, Holtman IR, Lerario AM, Vainchtein ID, Brouwer N, Sola PR, Veras MM, Pereira TF, Leite RE, Möller T, Wes PD, Sogayar MC, Laman JD, den Dunnen W, Pasqualucci CA, Oba-Shinjo SM, Boddeke EW, Marie SK, Eggen BJ. Transcriptomic analysis of purified human cortical microglia reveals age-associated changes. Nat Neurosci. 2017 Aug;20(8):1162-1171. Epub 2017 Jul 3 PubMed.
Srinivasan K, Friedman BA, Etxeberria A, Huntley MA, van der Brug MP, Foreman O, Paw JS, Modrusan Z, Beach TG, Serrano GE, Hansen DV. Alzheimer's Patient Microglia Exhibit Enhanced Aging and Unique Transcriptional Activation. Cell Rep. 2020 Jun 30;31(13):107843. PubMed.
Cantuti-Castelvetri L, Fitzner D, Bosch-Queralt M, Weil MT, Su M, Sen P, Ruhwedel T, Mitkovski M, Trendelenburg G, Lütjohann D, Möbius W, Simons M. Defective cholesterol clearance limits remyelination in the aged central nervous system. Science. 2018 Feb 9;359(6376):684-688. Epub 2018 Jan 4 PubMed.
Tcw J, Qian L, Pipalia NH, Chao MJ, Liang SA, Shi Y, Jain BR, Bertelsen SE, Kapoor M, Marcora E, Sikora E, Andrews EJ, Martini AC, Karch CM, Head E, Holtzman DM, Zhang B, Wang M, Maxfield FR, Poon WW, Goate AM. Cholesterol and matrisome pathways dysregulated in astrocytes and microglia. Cell. 2022 Jun 23;185(13):2213-2233.e25. PubMed. BioRxiv.
Make a Comment
To make a comment you must login or register.