Modern Microscopy Skims Surface of Living Minds and Spines
Quick Links
It is a safe bet that Anton van Leeuwenhoek and Robert Hooke, patriarchs of the light microscope, never saw this coming. Modern microscopy has taken their simple, light-bending lenses in new directions to create stunning images, including pictures of the living brain and detailed representations of the organ’s deep inner structure. New methods are emerging all the time to expand the toolbox of scientists working to understand neurodegeneration, and this series will highlight a few. For example, in the January 22 Nature Methods online, researchers from Cornell University in Ithaca, New York, describe how to mount a window over a mouse’s spinal cord so they can observe axon responses to injury over weeks or months. And in the February 3 Science, a team from the Max Planck Institute for Biophysical Chemistry in Göttingen, Germany, shows how nanometer-level resolution microscopy can pick out individual dendritic spines in a living mouse’s brain.
“When I was in medical school around 1970, light microscopy was considered a ‘mature’ technology,” joked Bill Betz of the University of Colorado School of Medicine in Denver. Turns out she was at best a teenager. In the early 1980s, the first confocal microscopes appeared, and things are still ‘maturing’ vigorously today, with the advent of optogenetics (reviewed in Fiala et al., 2010), super-resolution microscopy (reviewed in Lippincott-Schwartz and Manley, 2009), and more. Tech fans are further buoyed by a rainbow of fluorescent labels, such as quantum dots (see Park et al., 2012) for cell structures as well as pH and calcium ion reporters (see ARF related news story on Cheng et al., 2011; Ohki et al., 2005).
Several top research institutions, such as the Howard Hughes Medical Institute’s Janelia Farm in Ashburn, Virginia, and the Harvard Brain Center, are working to understand neural connections via optical microscopy, Betz said. “Optical imaging is probably the only method to give you non-invasive and high-resolution imaging,” said Chris Xu of Cornell University. Indeed, live imaging is a current trend in neurodegenerative research, said Jin-Moo Lee of Washington University in St. Louis, Missouri. A key advantage, he said, is that researchers can follow a single animal over time. In disease studies, traditional histology is like seeing a snapshot of an active battlefield and trying to answer the question “Who’s winning?” said Matthew Farrar of Cornell University, first author on the Nature Methods study. With longitudinal intravital studies, Lee said, researchers can watch how the battle unfolds, or watch how an intervention such as drug treatment affects neurons over time.
Through the Looking Glass
One common technique in intravital microscopy for neuroscience is to cut a window in the skull, or shave the skull until it is paper-thin, for two-photon imaging. Several teams have used such cranial windows to follow the kinetics of amyloid-β plaques in the brains of mouse models for Alzheimer’s disease (see Yan et al., 2009; Christie et al., 2001; ARF related news story; Prada et al., 2007; ARF news story) and tangles (ARF news story).
However, it has been challenging to open up a similar window over the spinal cord, where researchers might want to watch processes important to spinal cord injury or amyotrophic lateral sclerosis (ALS). Unlike the skull, the spinal cord bends and turns, and opening a vertebra exposes the animal to risk of spinal cord injury, said Chris Schaffer, senor author of the Nature Methods study. Moreover, the cord is located next to the ever-active heart and lungs, which could create motion artifacts in a microscopy image. Researchers have been able to longitudinally image the spinal cord by repeated surgeries (Dray et al., 2009), but that is hard on the animals, Schaffer said.
Farrar and Schaffer designed a procedure and equipment that sidestep those problems and enable them to watch axons in the spinal cord after injury. To see the spinal cord while keeping the spine safe from injury, Farrar surgically fused one thoracic vertebra to the ones immediately above and below. He then removed the dorsal lamina of the middle vertebra and installed a covering, like a mini-backpack with a clear window, above the opening. When the animals recovered from the procedure, they were able to move normally and he could image them at will. In most mice, fibrous tissue invading the viewing area blocked imaging after a couple of months. “Our ‘hero’ mouse went out just over five months,” Schaffer said. To avoid artifacts from breathing, the researchers designed their system so that the mouse was partially suspended by its spinal cord during imaging, such that any movement would not disturb the field of view.

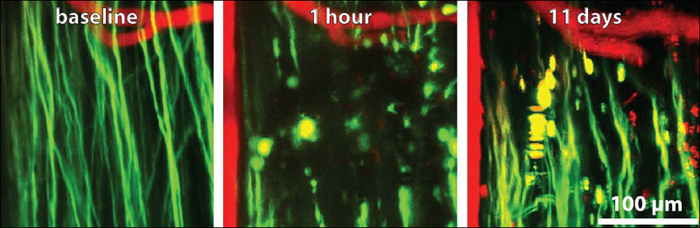
See the Spinal Cord Repair Itself
Using a chamber mounted over a mouse’s spinal cord and two-photon in-vivo microscopy, researchers observed nerve axons (green) and blood vessels (red) before axonal injury (left), one hour after (center), and 11 weeks after (right). (View larger image.) Image credit: Matthew Farrar, Cornell University, Ithaca, New York
Farrar used the system to watch how axons respond to laser-induced injury. He discovered that the response is highly heterogeneous: some axons died back within a day, others sprouted early on but then died, and some remained viable. Taken as a whole, the results were similar to other, single-time-point axon injury studies (Seif et al., 2007), but with the advantage of longitudinal imaging, the researchers were able to clearly see that individual axons respond differently to injury. The long-lasting “outliers” might be the best ones to approach therapeutically, Schaffer suggested, since they are already prone to survive.
“It is an exciting evolution of the [cranial window] technique to apply to multiple sclerosis, amyotrophic lateral sclerosis (ALS), or spastic parapesis,” as well as peripheral nerve injury, said Bradley Hyman of Massachusetts General Hospital in Boston, who was not involved in the study. For example, in ALS there has long been a question over whether glia or neurons are the first to exhibit pathology, Hyman said. Long-term imaging with Farrar’s chamber could help researchers answer that question.
Superior Resolution
For Stefan Hell and his colleagues at the Max Planck Institute, the authors of the Science paper, micron-level resolution is not enough. Hell developed a technique, called stimulated emission depletion (STED), to resolve two points to less than 70 nanometers with light microscopy—a feat previously thought to be impossible. The trick is to use two lasers. The first laser activates fluorophores. The second is a doughnut-shaped beam that quenches fluorophores on the edge of the activated area. In this way, STED minimizes the fluorescing area to a spot just a few nanometers across.
While Hell has used STED on nematodes before (Rankin et al., 2011), the research now reported in Science by first author Sebastian Berning and colleagues is the first example of using the technology in the brain of a living mammal, Hell said. Imaging through a glass cranial window, Berning was able to discern individual dendritic spines in six different mice. He saw that they appeared to move a bit on a timescale of minutes. Hell is not sure of the significance of these “slight relocations,” but neuroscientists think they may be important for learning, sensing, and memory formation (see ARF related news story on Trachtenberg et al., 2002 and Grutzendler et al., 2002). Berning was able to image 15-20 microns deep into the brain, but Hell believes that further advancements can push beyond that (Grotjohann et al., 2011).
“This super-resolution STED microscopy will allow us to examine protein localization and dynamics within tiny dendritic spines in the living animal, which was not possible to do in the past,” wrote Wenbiao Gan of New York University in an e-mail to ARF. “It would be interesting to see how β amyloid peptides and hyperphosphorylated tau are accumulated within the cell in living AD mice, and to see synaptic pathology over time at a more detailed level.” Gan was not part of the study team. While Hell has not yet performed longitudinal STED studies of in-vivo brains, he said it should be possible to do so.
For longitudinal imaging, researchers need to come back to the same place—the same neuron, plaque, or other structure—day after day or week after week. Typically, they use fluorescent dyes in blood vessels as markers, Hyman said. However, researchers in the laboratory of Mathias Jucker at the Hertie Institute in Tübingen, Germany, believe they have a more convenient system. Jasmin Hefendehl and colleagues super-glued a titanium ring atop the cranial window of their experimental mice. The ring has a groove that allows it to slide neatly onto the microscope stage. By programming the microscope to “remember” a given physical location, Hefendehl could return again and again to the same location for 25 weeks (Hefendehl et al., 2012). Other animals have worn the ring for a year and a half, she told ARF. Since she does not have to expend time and laser light looking for the reference blood vessels, there is less phototoxicity and photobleaching. “It really has made the experiments much easier,” Hefendehl said. She and Jucker have used the technique to follow amyloid plaque growth (see ARF related news story on Hefendehl et al., 2011).
Microscopy will mature further. In the future, scientists will have better techniques to regularly peer even more deeply into tissues of awake animals (e.g., see ARF related news story on Helmchen et al., 2001), Hyman predicted.—Amber Dance
References
News Citations
- Reel World—Viewing Long-Term Changes in Neuron Structure, Activity
- Popcorn Plaque? Alzheimer Disease Is Slow, Yet Plaque Growth Is Fast
- Caught in the Act—Amyloid Damages Neurons
- Imaging Tau and Caspases, Aβ’s Synaptic Effects
- Dendritic Spine Stability—Not So Black and White—or Is That Green and Yellow?
- Seeing Is Believing—Plaque Growth Is Slow, Tapers With Age
- Mini-Microscope Peers into Brains of Awake, Moving Rats
Paper Citations
- Fiala A, Suska A, Schlüter OM. Optogenetic approaches in neuroscience. Curr Biol. 2010 Oct 26;20(20):R897-903. PubMed.
- Lippincott-Schwartz J, Manley S. Putting super-resolution fluorescence microscopy to work. Nat Methods. 2009 Jan;6(1):21-3. PubMed.
- Park H, Li Y, Tsien RW. Influence of synaptic vesicle position on release probability and exocytotic fusion mode. Science. 2012 Mar 16;335(6074):1362-6. PubMed.
- Cheng A, Gonçalves JT, Golshani P, Arisaka K, Portera-Cailliau C. Simultaneous two-photon calcium imaging at different depths with spatiotemporal multiplexing. Nat Methods. 2011 Feb;8(2):139-42. PubMed.
- Ohki K, Chung S, Ch'ng YH, Kara P, Reid RC. Functional imaging with cellular resolution reveals precise micro-architecture in visual cortex. Nature. 2005 Feb 10;433(7026):597-603. PubMed.
- Yan P, Bero AW, Cirrito JR, Xiao Q, Hu X, Wang Y, Gonzales E, Holtzman DM, Lee JM. Characterizing the appearance and growth of amyloid plaques in APP/PS1 mice. J Neurosci. 2009 Aug 26;29(34):10706-14. PubMed.
- Christie RH, Bacskai BJ, Zipfel WR, Williams RM, Kajdasz ST, Webb WW, Hyman BT. Growth arrest of individual senile plaques in a model of Alzheimer's disease observed by in vivo multiphoton microscopy. J Neurosci. 2001 Feb 1;21(3):858-64. PubMed.
- Prada CM, Garcia-Alloza M, Betensky RA, Zhang-Nunes SX, Greenberg SM, Bacskai BJ, Frosch MP. Antibody-mediated clearance of amyloid-beta peptide from cerebral amyloid angiopathy revealed by quantitative in vivo imaging. J Neurosci. 2007 Feb 21;27(8):1973-80. PubMed.
- Dray C, Rougon G, Debarbieux F. Quantitative analysis by in vivo imaging of the dynamics of vascular and axonal networks in injured mouse spinal cord. Proc Natl Acad Sci U S A. 2009 Jun 9;106(23):9459-64. PubMed.
- Seif GI, Nomura H, Tator CH. Retrograde axonal degeneration "dieback" in the corticospinal tract after transection injury of the rat spinal cord: a confocal microscopy study. J Neurotrauma. 2007 Sep;24(9):1513-28. PubMed.
- Rankin BR, Moneron G, Wurm CA, Nelson JC, Walter A, Schwarzer D, Schroeder J, Colón-Ramos DA, Hell SW. Nanoscopy in a living multicellular organism expressing GFP. Biophys J. 2011 Jun 22;100(12):L63-5. PubMed.
- Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature. 2002 Dec 19-26;420(6917):788-94. PubMed.
- Grutzendler J, Kasthuri N, Gan WB. Long-term dendritic spine stability in the adult cortex. Nature. 2002 Dec 19-26;420(6917):812-6. PubMed.
- Grotjohann T, Testa I, Leutenegger M, Bock H, Urban NT, Lavoie-Cardinal F, Willig KI, Eggeling C, Jakobs S, Hell SW. Diffraction-unlimited all-optical imaging and writing with a photochromic GFP. Nature. 2011 Oct 13;478(7368):204-8. PubMed.
- Hefendehl JK, Milford D, Eicke D, Wegenast-Braun BM, Calhoun ME, Grathwohl SA, Jucker M, Liebig C. Repeatable target localization for long-term in vivo imaging of mice with 2-photon microscopy. J Neurosci Methods. 2012 Apr 15;205(2):357-63. PubMed.
- Hefendehl JK, Wegenast-Braun BM, Liebig C, Eicke D, Milford D, Calhoun ME, Kohsaka S, Eichner M, Jucker M. Long-term in vivo imaging of β-amyloid plaque appearance and growth in a mouse model of cerebral β-amyloidosis. J Neurosci. 2011 Jan 12;31(2):624-9. PubMed.
- Helmchen F, Fee MS, Tank DW, Denk W. A miniature head-mounted two-photon microscope. high-resolution brain imaging in freely moving animals. Neuron. 2001 Sep 27;31(6):903-12. PubMed.
Other Citations
{kind=link}
Further Reading
Papers
- Misgeld T, Kerschensteiner M. In vivo imaging of the diseased nervous system. Nat Rev Neurosci. 2006 Jun;7(6):449-63. PubMed.
- Scheibe S, Dorostkar MM, Seebacher C, Uhl R, Lison F, Herms J. 4D in in vivo 2-photon laser scanning fluorescence microscopy with sample motion in 6 degrees of freedom. J Neurosci Methods. 2011 Aug 30;200(1):47-53. PubMed.
- Dibaj P, Steffens H, Zschüntzsch J, Kirchhoff F, Schomburg ED, Neusch C. In vivo imaging reveals rapid morphological reactions of astrocytes towards focal lesions in an ALS mouse model. Neurosci Lett. 2011 Jun 22;497(2):148-51. PubMed.
- Kaminski Schierle GS, van de Linde S, Erdelyi M, Esbjörner EK, Klein T, Rees E, Bertoncini CW, Dobson CM, Sauer M, Kaminski CF. In situ measurements of the formation and morphology of intracellular β-amyloid fibrils by super-resolution fluorescence imaging. J Am Chem Soc. 2011 Aug 24;133(33):12902-5. PubMed.
- Hadley KC, Borrelli MJ, Lepock JR, McLaurin J, Croul SE, Guha A, Chakrabartty A. Multiphoton ANS fluorescence microscopy as an in vivo sensor for protein misfolding stress. Cell Stress Chaperones. 2011 Sep;16(5):549-61. PubMed.
- Barretto RP, Ko TH, Jung JC, Wang TJ, Capps G, Waters AC, Ziv Y, Attardo A, Recht L, Schnitzer MJ. Time-lapse imaging of disease progression in deep brain areas using fluorescence microendoscopy. Nat Med. 2011 Feb;17(2):223-8. PubMed.
- Drew PJ, Shih AY, Driscoll JD, Knutsen PM, Blinder P, Davalos D, Akassoglou K, Tsai PS, Kleinfeld D. Chronic optical access through a polished and reinforced thinned skull. Nat Methods. 2010 Dec;7(12):981-4. PubMed.
News
- Kiss and Tell—STED Microscopy Resolves Vesicle Recycling Question
- Window to the Brain Shows Dystrophic Neurites Shrinking
- Philadelphia: Diagnosis Light—Toward Infrared Plaques
- Reel World—Viewing Long-Term Changes in Neuron Structure, Activity
- Photons Light up Neuronal Circuits, Pinpoint Single Cells
- Popcorn Plaque? Alzheimer Disease Is Slow, Yet Plaque Growth Is Fast
- Caught in the Act—Amyloid Damages Neurons
- Imaging Tau and Caspases, Aβ’s Synaptic Effects
- Dendritic Spine Stability—Not So Black and White—or Is That Green and Yellow?
- Seeing Is Believing—Plaque Growth Is Slow, Tapers With Age
- Mini-Microscope Peers into Brains of Awake, Moving Rats
Primary Papers
- Farrar MJ, Bernstein IM, Schlafer DH, Cleland TA, Fetcho JR, Schaffer CB. Chronic in vivo imaging in the mouse spinal cord using an implanted chamber. Nat Methods. 2012 Mar;9(3):297-302. PubMed.
- Berning S, Willig KI, Steffens H, Dibaj P, Hell SW. Nanoscopy in a living mouse brain. Science. 2012 Feb 3;335(6068):551. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.